극피동물
다른 표기 언어 Echinoderm요약 해산 무척추동물 가운데 한 문인 극피동물문에 속하는 동물들.
목차
펼치기-
특징
- ┗ 크기와 구조의 다양성
- ┗ 분포와 수
- ┗ 자연계에서의 역할
- ┗ 인간과의 관계
- 생식과 생활사
-
먹이와 섭식
- 운동
- 생태
-
형태와 기능
-
외부형태
- ┗ 대칭과 체형(體形)
- ┗ 골격
- 내부형태
- 진화
- 분류
-
한국의 극피동물
21강으로 나뉘고, 현존하는 것으로는 바다나리강·성게강·해삼강·불가사리강·거미불가사리강과 최근에 발견된 콘켄트리키클로이데아강이 있다.
약 5억 7,000만 년 전 캄브리아기 전기에 시작된 동물로 풍부한 화석이 나오나 그 대부분은 현재 멸종되었다.
19세기초, 별개의 동물군으로 인식되어 있던 극피동물은 때로 자포동물과 함께 방사상칭성동물로 묶이기도 했다. 현존하는 것이 6,000종 이상, 화석종이 1만 3,000종 이상 알려져 있다.
특징
크기와 구조의 다양성
극피동물 대부분은 크기가 10cm 이하로 작으나 상당히 큰 것도 있다. 해삼류 중 어떤 것은 2m, 일부 불가사리류는 1m까지 달하며, 가장 큰 극피동물은 멸종된 갯고사리류로 약 20m에 이른다.
극피동물은 체형에서 다양함을 보이는데 멸종된 군에서 더욱 특이하다. 병부(柄部)를 가진 꽃과 같은 갯고사리류에서 벌레 모양의 해삼류, 딱딱한 불가사리류와 성게류에 이르기까지 모양이 다양하다.
또한 빨강·노랑·초록·자주색과 같은 매우 아름다운 색깔을 띤다. 열대에 분포하는 종은 어두운 갈색에서 검정색을 띠기도 하나, 강한 태양에 노출되지 않은 종들은 밝은 색깔 특히 노란색을 띤다.
분포와 수
극피동물은 전세계에서 발견된다. 보통 성게류와 불가사리류는 연안에 분포한다. 대부분은 특히 온대지역에 국한되어 인도양과 태평양에 널리 분포하나 일부 북극과 남극, 또 열대종은 그 분포구역이 매우 넓다.
또한 수직적 분포도 매우 제한되어 있다. 즉 연안에 분포하는 종은 100m 이상의 깊이에는 있지 않다. 일부 심해종은 1,000m에서 5,000m 이상까지 상당히 깊은 곳에서도 발견된다.
자연계에서의 역할
극피동물은 해저의 부패물질을 먹는 매우 효율적인 부식성 동물이기도 하며, 많은 양의 작은 생물들을 먹어 그들의 수를 조절하기도 한다. 또한 해저 침전물의 구조를 여러 방법으로 바꾸기도 한다. 한 지역의 큰 해삼류 집단은 해저 침전물의 물리적·화학적 구성을 크게 변화시킬 수도 있다.
그뿐 아니라 극피동물은 많은 수의 유생을 만들어 다른 플랑크톤의 먹이가 되기도 한다(→ 수중생태계).
인간과의 관계
열대산 일부 대형 해삼류는 상업적으로 갈이(trepang)로 알려져 있으며, 특히 아시아에서는 말려서 국으로 먹는다. 성게류의 생식선은 유럽·지중해·일본·칠레 지역에서 진미로 여겨지고 있다. 일부 열대산 해삼류는 많은 동물을 죽이는 홀로투린(holothurin)이라고 알려진 독소를 갖고 있으나, 인체에는 해롭지 않은 것 같다.
사실상 독소는 종양의 성장률을 줄이므로 의학적으로 중요한 것으로 알려져 있다. 특히 성게류와 불가사리류의 생식소는 쉽게 얻을 수 있어서 발생학 연구에 사용되기도 한다. 캘리포니아 연안을 따라 있는 성게류는 어린 해산식물을 먹어 상업상 중요한 해조의 성장을 저해하기도 한다.
생식과 생활사
개요
대부분의 극피동물은 자웅이체로, 보통 유성생식을 한다.
일부 해삼류·불가사리류·거미불가사리류는 무성생식을 하기도 한다.
무성생식
몸이 둘 또는 그 이상으로 나누어지는 분열(fission)과 잃어버린 몸 부분을 다시 만들어내는 재생(regeneration)이 있다.
분열은 약 60여 종의 불가사리류와 거미불가사리류, 해삼류에서 흔하게 일어나는 생식방법으로 현존 종의 약 1%에 해당하며, 이들 중 어떤 것의 유성생식은 알려져 있지 않다.
재생력은 특히 바다나리류와 불가사리류, 거미불가사리류에서 잘 알려져 있다. 불가사리를 잡아 찢은 다음 바다로 버릴 경우 더 많은 수로 불어난다. 일부 해삼류는 공격을 받거나 환경이 좋지 않은 경우 또는 계절적 이유로 그들의 내장기관을 내뿜어버리고(탈내장), 몇 주 안에 재생한다.
성게류도 잃어버린 내부 골격의 작은 부분이나 차극(pedicellariae), 또는 극 등을 쉽게 재생할 수 있다.
유성생식
방출암컷에서 수백만 개의 난자와 수컷에서 정자가 바다로 방출(산란)된 다음 수정된다.
대부분의 극피동물은 1년마다 산란하는데, 산란시기는 보통 봄 또는 여름이고, 1~2개월 동안 지속된다. 그러나 어떤 종은 1년 내내 산란하기도 한다. 산란을 유도하는 원인은 복잡하며, 온도·빛·해수의 염분도와 같은 외적 영향을 받는 것 같다. 대부분의 극피동물은 산란 전에 집합해서 수정률을 높인다.
또한 일부는 산란 동안 특징있는 행동을 하기도 한다.
일부 불가사리류와 거미불가사리류는 몸의 중앙 부위를 해저의 바닥으로부터 들어올리며, 해삼류는 몸의 앞쪽 끝을 올려서 흔들기도 한다. 이러한 움직임은 아마 난자와 정자가 해저 침전물 속에 박히는 것을 막기 위함인 것 같다.
발생
수정난은 여러 방식으로 배발생을 한다.
난황이 적은 작은 알은, 작은 생물을 먹고 사는 플랑크톤인 자유유영하는 유생으로 발달한다. 난황이 많은 더 큰 알은 플랑크톤인 유생형으로 발생하나 작은 생물을 먹기보다는 갖고 있는 난황을 섭취한다. 알, 플랑크톤인 유생단계, 어린 극피동물로의 발생을 간접발생이라고 한다. 많은 난황을 갖고 있는 큰 알들은 유생단계를 거치지 않는데 이를 직접발생이라고 한다. 직접발생에서는 어린 것들이 보통 어미의 보호를 받는다.
육낭보호(brood protection)는 남극과 북극, 심해산 극피동물에서 가장 잘 알려져 있다.
간접발생에서 수정난은 여러 번 분열하여 포배단계의 섬모성 세포군을 만들어낸다. 난할은 전할(全割), 부정분할(不定分割) 그리고 방사난할(放射卵割)을 한다. 포배는 계속해서 낭배가 된다. 극피동물은 후구동물에 속하는 척추동물과 일부 무척추동물(모악동물과 반색동물)에 유사한 발생과정을 갖는다.
낭배는 극피동물의 기본 유생형인 좌우대칭성 디플류룰라(dipleurula)가 된다.
해삼류의 디플류룰라는 아우리쿨라리아(auricularia)로 되며 불가사리류는 비핀나리아(bipinnaria)에서 브라키올라리아(brachiolaria)로 된다.
성게류의 에키노플루튜스(echinopluteus)와 거미불가사리류의 오피오플루튜스(ophiopluteus)는 복잡하고 진보된 유생형으로 형태상 유사하다.
갯고사리는 디플류룰라 단계가 없이 돌리올라리아로 된다.
유생형은 대부분 작고 가끔 길이가 1㎜ 이하이지만 일부 해삼류는 15㎜, 불가사리류의 비핀나리아는 25㎜에 달하기도 한다. 자유유영하는 플랑크톤으로 수일이나 수주를 지낸 유생은 복잡한 변태(變態) 과정을 겪은 다음 어린 극피동물이 되며, 변태 동안 기본적인 좌우대칭은 방사대칭으로 되어 5개의 수관을 형성하게 된다.
극피동물의 평균 수명은 약 4년이고 일부 종은 8~10년까지도 산다.
먹이와 섭식
극피동물은 다양한 방법으로 먹이를 먹는다. 분명한 섭식 리듬이 있어서 대부분은 밤에만 먹지만 어떤 것들은 계속해서 먹는다. 식성은 활발하며, 선택적인 육식성에서부터 잡식성인 부식성 또는 무작위적으로 진흙을 삼키는 것까지 매우 다양하다.
바다나리류는 현탁물(懸濁物)을 먹는데, 촉수나 완(腕) 위의 홈에 있는 관족이 만든 점액망으로 플랑크톤을 잡아먹는다. 대부분의 불가사리류는 활발한 육식자로서 조개류 또는 다른 불가사리류를 잡아먹는다. 일부 종은 진흙을 삼키기도 한다.
일부 불가사리류는 입을 통해 먹이로 위(胃)를 밀어내 외부에서 먹이를 일부 소화시킨 뒤, 위를 다시 몸안으로 넣어 몸안에서 먹이를 완전히 소화시킨다. 대부분의 거미불가사리류는 바다에 떠다니거나 바닥에 놓인 작은 생물을 먹는데, 완과 관족으로 잡아서 입까지 보낸다. 완이 복잡하게 가지를 치고 있는 거미불가사리류는 바다나리류와 비슷하게 먹는다. 콘켄트리키클로이데아강의 섭식방법은 아직 알려져 있지 않다.
원시형인 소위 정형(正形) 성게류는 잡식성이거나 채식성으로, 바위에 있는 해조류나 작은 생물들을 그들의 단단한 이로 갉아 먹는다. 좀더 발달된 형인 부정형(不正形) 성게류는 이가 없으므로 해저 침전물을 파고들어 극과 관족으로 작은 먹이를 입까지 운반한다. 일부 해삼류는 표면에 붙어 있으면서, 가지가 쳐진 끈끈한 촉수들의 망으로 플랑크톤을 잡아먹고, 일부는 해저면에서 촉수로 먹이를 잡아 입에 넣는다. 대부분의 해삼류는 육상 지렁이처럼 진흙·모래를 삼켜서 유기물질을 소화하고 노폐물을 내보낸다.
운동
개요
불가사리류와 성게류는 극과 관족을 사용하여 움직인다. 콘켄트리키클로이데아강도 아마 같은 방법으로 움직이는 것 같다. 거미불가사리류는 어떤 식으로든 완을 휘둘러 움직인다. 그 가운데는 배를 젓는 듯한 움직임도 있어 2쌍의 완으로 노를 젓듯 하는데, 다섯번째 완은 앞쪽으로 나가거나 뒤따른다. 해삼류는 보통 입 쪽이 앞서며 관족과 몸의 수축·이완으로 움직인다.
유병(有柄) 바다나리류, 즉 바다술류는 보통 부착기라고 불리는 병부의 끝에 있는 구조물로 표면에 단단히 붙어 있으며, 새로운 부착장소를 찾아 움직이기도 한다. 무병 바다나리류, 즉 깃별나리류는 보통 그들의 수많은 완을 위아래로 움직여 유영한다.
유영은 바다나리류와 거미불가사리류, 해삼류에서 알려져 있다.
바로서기 반응
극피동물 가운데 입이 표면을 향하는 것은 불가사리류, 거미불가사리류, 콘켄트리키클로이데아강 동물들, 성게류이고 입이 표면에서 멀리 있는 것은 바다나리류와 해삼류이다. 뒤집혔을 때 극피동물은 바로 서는 반응을 보인다. 불가사리류는 관족을 사용하여 가장 효과적으로 반응한다. 성게류도 관족과 극을 사용하여 몸을 바로잡는다. 해삼류·바다나리류·거미불가사리류와 같이 민첩한 극피동물은 비교적 쉽게 바로잡는 반응을 한다.
굴파기
많은 극피동물은 바위나 부드러운 저질 속에 굴을 파고 산다. 갯고사리류는 그들의 섭식기구가 깨끗하게 치워져야 하므로 굴을 파지 않는다. 일부 성게류는 이와 극을 사용하여 바위 속을 3cm 이상씩 파고 산다. 소위 부정형 성게류는 부드러운 저질 밑에 들어가서 사는데 염통성게류는 해저 38cm 깊이에서도 발견된다. 해삼류는 촉수와 체벽의 수축으로 굴을 판다. 불가사리류도 모래나 진흙 속에 들어가 살기도 한다. 일부 거미불가사리류는 먹이를 잡기 위해 완의 끝만을 밖에 내놓고 해저면 밑에 들어가 산다.
생태
서식처
극피동물은 모두 해산이며, 단지 몇 종만이 기수지역에도 있다. 대부분의 종은 염도와 온도, 빛의 세기에 현저한 변화가 있으면 이것을 견뎌내지 못하고 다른 곳으로 이동한다. 천해에 사는 종들 대부분은 행동이 빛에 의해 조절된다. 또한 대다수 종의 수평·수직 분포는 해수의 온도에 의해 결정된다. 압력의 영향은 아직 조사되지 않았다.
극피동물은 서식처가 다양하다. 암석 해안을 따라 불가사리류와 성게류가 바위에 붙어 있으며, 그 바위 밑에는 해삼류와 거미불가사리류가 숨어 있다. 모래지역에서는 불가사리류와 거미불가사리류, 부정형 성게류, 해삼류가 해저면 밑이나 위에 살고 있다. 현존 극피동물군의 큰 집단은 진흙이나 펄 지역에서 볼 수 있다. 일부 해양에서는 극피동물이 우점하고 있는 경우도 있다. 예를 들면, 심해에서는 해삼류가 현존량의 90% 이상을 차지하기도 한다.
극피동물은 빈번하게 다른 동물을 집으로 사용한다. 예를 들면, 수천 마리의 거미불가사리류가 어떤 열대산 해면동물 속에 살고 있는 경우도 있다. 해삼류가 남극 성게류의 극에 붙어 있는 경우가 있으며, 어떤 해삼은 심해산 어류의 피부에 붙어 산다. 반면에 극피동물은 많은 생물의 숙주가 되기도 한다. 예를 들면, 여러 가지 갑각류와 따개비류는 성게류의 골격에 충영을 이루면서 살고, 바다나리류는 특이한 기생성 벌레의 숙주이기도 하다. 여러 열대산 해삼류와 해삼류의 직장에서 발견되는 숨이고기는 흥미있는 공생관계를 나타내기도 한다.
육식과 방어
일반적으로 극피동물집단은 다른 동물에 잡아먹히지 않으나, 거미불가사리류는 여러 어류와 일부 불가사리류의 중요한 먹이원이 되기도 한다. 성게는 자주 판새류·경골어류·물맞이게류·복족류에 먹히며, 까마귀·재갈매기·아이더오리류·북극여우·라코·사람이 대량으로 먹기도 한다. 불가사리류는 다른 불가사리류나 연체동물, 갑각류에 먹힌다. 일부 해삼류는 어류와 사람이 잡아먹는다. 바다나리류는 일정한 포식자가 없는 것 같다.
극피동물은 여러 수동적인 방법으로 자신을 보호한다. 딱딱한 골격은 포식자를 저지한다. 예를 들면, 성게류는 극(棘)과 독이 있는 차극을 갖고 있다. 일부 불가사리류는 화학물질을 분비해서 포식성 연체동물을 피하기도 한다. 몇몇 해삼류는 호흡수로 알려진 끈끈한 흰 실덩이를 항문 밖으로 방출하고, 일부는 독소인 홀로투린을 지니고 있다.
집합
극피동물은 많은 수로 모이는 경향이 있는데, 과거에도 역시 그랬던 것 같다. 현재 거미불가사리류는 해저의 많은 부분을 덮고 있으며, 성게류도 흔히 널리 모여 있다. 해삼류와 바다나리류, 일부 불가사리류도 흔히 집합하는 경향을 보인다.
집합 현상은 먹이를 얻을 수 있는가 하는 것 등 여러 환경요인에 대한 반응이다. 즉 거미불가사리류나 바다나리류 대부분은 플랑크톤의 양이 많은 강한 해류가 흐르는 지역을 점유하고 있다. 거미불가사리류는 서로를 지탱하면서 완을 물 속으로 내어 먹이를 잡는다. 집합은 개체들 사이의 상호작용의 결과물인 사회적 행동을 확실히 보여주는 것일 뿐 아니라, 개체들과 환경의 상호작용 결과 생긴 수동적 현상인 것 같다.
형태와 기능
극피동물은 작고 수많은 석회성(탄산칼슘) 판들로 이루어진 골격을 가진다. 체강은 수관계로서 관족과 피새, 그리고 운동·섭식·호흡·감각지각에 사용되는 구조물처럼, 체표면에 돌출한 체액으로 채워진 관들로 이루어져 있다.
외부형태
대칭과 체형(體形)
초기 극피동물 대부분은 대칭성이 없거나 좌우대칭이었다.
좌우대칭은 모든 현생종에서 나타나며, 유생단계에 특히 뚜렷하다. 오방사(五放射) 대칭은 현존 극피동물의 모든 군에서 뚜렷하며 관족의 배열에서 분명해진다. 수관계·근육계·혈관계·신경계 또한 오방사형이다. 골격도 현미경적 골편으로 퇴축된 해삼류를 제외하고는 오방사형이다.
극피동물은 서로 다른 성장형에 따라 여러 아문(亞門)으로 나뉜다.
호말로조아아문(Homalozoa)은 비대칭형이고, 바다나리아문(Crinozoa)·블라스토조아아문은 입으로부터 관족이 악부를 가로질러 완까지 뻗어 있으며, 불가사리아문(Asterozoa)은 관족이 입으로부터 완까지 퍼져 있고 별 모양을 이룬다. 성게아문(Echinozoa)에서는 관족이 구형의 몸에 5개의 줄을 이루고 있다. 바다나리류는 입이 컵 모양의 악부 중앙에 있고 많은 수의 완이 악부에 붙어서 고사리의 잎과 비슷해 보인다.
완은 기본적으로 5개이지만 대부분의 현존 종에서는 한 번 또는 여러 번 가지를 쳐서 10~200개에 달하기도 한다. 악부 밑에 붙어 있는 병부를 가지거나, 대부분의 현존 종에서 보듯이 병부가 없이 권수(cirri)로 부착한다.
불가사리류는 중앙에 커다란 반(disk)이 있으며, 그곳으로부터 중요한 내부 기관계 일부를 담고 있는 속이 빈 완이 5개나 그 이상 뻗어 있다. 반의 아랫면 중앙에 입이 있고, 각 완의 아랫면 보대구(ambulacral furrow)라고 불리는 특별한 홈 속에 5개 또는 그 이상의 관족 열(列)이 있다.
반의 윗면 중앙에는 항문(가끔 없기도 함)과 천공판(madreporite)이 있다. 불가사리 가운데는 7개의 완을 갖는 것이 많이 있으며, 한 심해산과는 6~20개의 완을 가지고 있고, 남극해(海)에 사는 한 속은 완을 50개까지 가진다.
콘켄트리키클로이데아강 동물들은 둥근 쟁반 모양으로 등쪽 표면은 판으로 되어 있으나 배쪽 표면은 노출되어 있다.
거미불가사리류는 작은 반으로부터 5개의 완이 방사상으로 나 있으며 커다란 내부기관들은 보통 반에 국한되어 있다.
입은 특별한 홈에 들어 있지 않은 관족처럼 반의 아랫면 중앙에 있다. 대부분의 거미불가사리류는 5개의 완을 가지나, 일부 종은 6개 또는 그 이상의 완을 갖기도 하고, 삼처발이류에서는 5개의 완이 가지를 쳐 복잡한 망(網) 모양을 이룬다.
성게류는 골격이 단단한 판으로 이루어진 둥근 껍데기를 이룬다. 관족은 판의 구멍으로 나오고 5개의 눈에 잘 띄는 띠, 즉 보대(步帶)를 이루며 보대 사이를 간보대라고 한다. 정형 성게류는 대략 구형이며, 입은 5개의 보대가 만나는 중앙에 있고, 항문은 입의 반대쪽에 있다.
부정형 성게류는 길거나 납작하여 항문이 입쪽 또는 반대쪽에 있다. 정형과 일부 부정형 성게류는 이가 5개 있으며, 아리스토틀등(燈)이라고 하는 판과 근육으로 된 복잡한 계(系)가 이를 조종한다.
해삼류는 몸이 길고, 입과 항문은 서로 반대쪽에 있다. 관족은 몸의 아랫면에 더 많이 있거나 또는 없다. 바깥쪽은 좌우대칭이고 안쪽은 방사대칭이다.
골격
골격은 진피성(眞皮性)이나 해삼류를 제외하고는 뚜렷하며, 효과적으로 갑옷을 이루고 있다.
각 골격단위(골편)는 진피층에서 과립으로부터 만들어지는데, 과립이 특수한 석회분비세포에서 분비된 후 합쳐진다. 골격의 부분들은 동물이 자라면서 커지며, 골격의 재흡수와 재생이 일어나기도 한다.
극피동물은 골격구조가 다양하다. 성게류의 골격은 판으로 된 10개의 원주로 이루어져 있으며, 방어 또는 청소에 사용되는 차극뿐 아니라 크고 작은 극을 갖고 있다. 복잡한 섭식기구인 아리스토틀등은 근육과 결합조직에 의해 붙어 있는 40개의 골편으로 되어 있다. 바다나리류는 각각 5개의 골격판으로 이루어진 속이 빈 악부로 되어 있으며, 병부와 권수는 드럼통 모양의 골편으로 이루어져 있다.
불가사리류의 골격은 수많은 극을 갖는 여러 모양의 골편으로 되어 있으나 근육과 인대로 붙어 있어 유연하다. 완은 중앙축이 척추라 불리는 긴 골편으로 이루어져 있다.
해삼류에서는 현미경적 크기의 골편이 구멍이 있는 납작한 방패판 모양에서부터 방사성의 수레 모양까지 다양한 모양을 하고 있으며 그 수도 많다. 예를 들면 한 열대종은 2,600만 개의 골편을 갖고 있다. 석회환(calcareous ring)이라 불리는 판이 입에서 위까지 이르는 관을 둘러싸고 있으며, 이는 섭식기구가 아니라 근육의 삽입점으로서 기능한다.
내부형태

수관계(水管系)
수관계는 극피동물의 특징적 구조물로, 관족을 움직이게 한다.
수액을 갖고 있어 관들의 내부수압계를 이루며, 천공판과 석관이 연결되어 있는 환상수관(環狀水管)·방사수관(放射水管)으로 이루어져 있다. 천공판은 보통 바깥에 있으며 물을 받아들인다. 물은 환상수관을 거쳐 방사수관을 따라 관족에 이르며, 관족은 방사관에 있는 근육이나 병낭(ampulla)의 수축에 의해서 움직인다.
수관계의 구조는 분류군에 따라 다양하다.
천공판은 불가사리류의 경우 보통 1개 이상 있고, 해삼류에서는 보통 체강 내에 있다. 관족은 붙는 힘이 강한 잘 발달한 흡반을 갖고 있고 뾰족하며 호흡·섭식·굴파기·점액 생산 또는 감각지각을 하도록 적응되었다.
체벽과 체강
거미불가사리류를 제외한 대부분의 극피동물은 표피에 섬모가 있는데 바다나리류의 체벽에는 비교적 적게 있다.
섬모는 입까지 먹이 입자를 옮기거나 청소를 한다. 또한 샘[腺]세포와 감각세포도 표피에 있다. 극과 차극 같은 골격성분의 표피 아래층은 진피로서 석회성 골격과 결합조직을 포함하고 있다. 진피의 안쪽에는 횡근과 종근이 있다. 체강은 위장강과 수관계 2부분으로 나뉘며, 여러 특별한 부분을 만들고 있다.
소화계와 혈관계
소화관은 하나의 관으로 이루어져 있는데 곧거나(불가사리류와 거미불가사리류), 시계 방향으로 감겨 있거나(바다나리류와 해삼류), 처음에는 시계 방향으로 감겼다가 나중에 시계 반대 방향으로 감겨 있다(성게류). 관은 식도·위·장 그리고 직장으로 나뉜다.
항문은 거미불가사리류와 일부 불가사리류에는 없으나, 대부분의 극피동물에 있다(소화관).
혈관계는 체강의 일부도 진정한 혈관도 아닌 중간의 복잡한 계를 이루고 있다. 환상혈관과 5개의 방사혈관들은 식도와 수관계의 방사관을 둘러싸고 있다. 복잡한 혈관망은 소화관과 생식소와 연관되어 있다.
축기관
축기관은 해삼류를 제외한 모든 극피동물에서 볼 수 있으며, 복잡하고 긴 조직덩어리로, 위장강·수관계·혈관계의 공통 연결부위이다.
이것은 기능이 아직 밝혀져 있지 않으나 공격 생물에 대해 방어기능을 하며, 수축할 수 있어 체액의 순환을 일으키고 배설·분비활동도 하는 것 같다.
신경계와 감각기관
극피동물은 신경계가 복잡하다.
피부 아래에 신경총(叢)이 하나 있고, 1개에서 여러 개의 신경환이 식도를 둘러싸고 있으며 이곳으로부터 흔히 수관계와 평행하게 방사신경이 나온다. 환상신경과 방사신경은 몸의 바로잡기에 관계한다.
극피동물은 소수의 잘 발달된 감각기관을 갖지만 그들은 감촉·광도·온도·방향·해수의 변화에 민감하다. 관족·극·차극·피부는 촉각으로 반응하며, 빛을 감지하는 기관은 성게류·해삼류·불가사리류에서 볼 수 있다.
생식계
바다나리류는 생식소(生殖巢)가 완과 우지에 있는 특별한 강 내에 있어, 주요 체강의 밖에 생식소가 있는 유일한 극피동물이다.
불가사리류는 각 완의 기부에 2개씩, 10개의 생식소가 있다. 콘켄트리키클로이데아강 동물들은 주머니 같은 생식소 5쌍을 갖고 있다. 거미불가사리류는 생식소가 체강에 매달려 있는 주머니에 붙어 있다. 주머니는 완의 기부에서 몸 밖으로 열리며, 각각에 1~1,000여 개까지의 생식소가 붙어 있다. 정형 성게류의 경우 보통 간보대에 5개의 생식소가 붙어 있다.
생식소관은 항문 가까이에서 밖으로 열린다. 대부분의 부정형 성게류는 4개의 생식소를 갖고 있으나, 일부 종은 3개 또는 5개, 몇 종은 2개의 생식소를 갖고 있다. 해삼류는 다른 극피동물과 달리 1개의 생식소를 가지며, 몸의 앞쪽 끝에서 밖으로 열려 있다.
진화
멸종 극피동물
극피동물이 캄브리아기 전기에 이미 아주 다양했던 것을 보면, 선캄브리아시대에 상당한 진화가 일어난 것이 틀림없다.
오스트레일리아에서 발견된 선캄브리아시대의 화석은 삼방사대칭이며 외관상 좌해성류와 유사하다. 이것은 삼방사대칭형이 오방사대칭의 선구형이라는 것과 이 화석이 원시극피동물인 것을 암시하고 있다.
과학자들은 선캄브리아시대 극피동물의 화석이 없는 것은 초기 극피동물이 수관계는 갖고 있었을지 모르나, 석회성 골격은 없어서 화석화되지 않았다는 것을 나타낸다고 생각한다.
현존 극피동물
현존 극피동물을 구성하는 여러 강 사이의 유연관계에 대해서는 수십 년 동안 논쟁이 있었다. 일부 과학자는 유생단계가 군들의 상호관계를 반영한다고 믿는다. 따라서 성게류와 거미불가사리류를 한 군으로, 불가사리류와 해삼류를 한 군으로 본다. 몇 가지 생화학적인 연구도 이러한 안을 뒷받침한다.
반면에 비교해부학과 일부 고생물학 연구에 따르면, 거미불가사리류와 불가사리류는 아마도 바다나리류 비슷한 조상에서 생겨났다고 보고 있다.
분류
개요
극피동물의 분류는 1970~80년대에 대변동을 겪었으며 아직도 논쟁중에 있다.
어느 누구도 분류체계에 만족하지 못하나 극피동물의 경우는 더욱 그렇고 새로운 발견과 새로운 이론이 계속해서 첨가되고 있다. 여기에서는 5문으로 나누었는데, 고생물학자와 동물학자들에 의한 최근의 연구를 근거로 하였다. 아문 준위 아래에서 분류기준은 다양하나, 가장 중요한 특성을 골격에 두었다. 멸종되어 화석으로만 알려져 있는 강은 ¶로 표시하였다.
극피동물문(棘皮動物門 Echinodermata)은 전세계에 널리 분포하는 해산 무척추동물로 오방사대칭을 나타내며, 골격은 탄산칼슘(CaCO3)으로 구성되어 있다.
체강은 수관계를 이루고 있으며, 캄브리아기부터 현재까지 있으며, 약 1만 3,000종의 멸종종과 6,000여 종의 현생종이 있다.
호말로조아아문(Homalozoa)
캄브리아기 중기~데본기 중기(약 3억 6,500만~5억 7,000만 년 전).
①¶스틸로포라강(Stylophora) : 캄브리아기 중기~오르도비스 후기(약 4억 6,000만~5억 4,000만 년 전).
②¶호모스텔레아강(Homostelea) : 캄브리아기 중기(약 5억 4,000만 년 전).
③¶호모이오스텔레아강(Homoiostelea) : 캄브리아기 후기부터 데본기 전기까지(약 4억~5억 1,000만 년 전).
④¶크테노키스토이데아강(Ctenocystoidea) : 캄브리아기 중기(약 5억 4,000만 년 전).
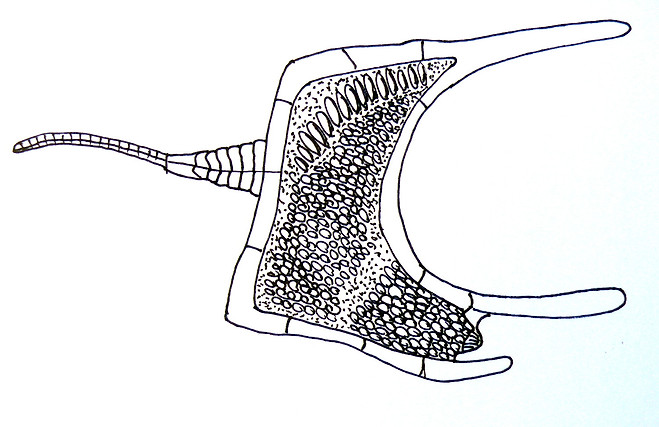
블라스토조아아문(Blastozoa)
캄브리아기~페름기(약 2억 8,000만~5억 4,000만 년 전).
①¶에오크리노이데아강(Eocrinoidea) : 캄브리아기 전기~실루리아기(약 4억 3,000만~5억 7,000만 년 전).
②¶블라스토이데아강(Blastoidea) : 실루리아기~페름기(약 2억 8,000만~4억 3,000만 년 전).
③¶파라크리노이데아강(Paracrinoidea) : 오르도비스기 중기(약 4억 6,000만 년 전).
④¶파라블라스토이데아강(Parablastoidea) : 오르도비스기 전기~중기(약 4억 6,000만~5억 년 전).
⑤ ¶롬비페라강(Rhombifera) : 오르도비스기 전기~데본기 후기(약 3억 5,000만~5억 년 전).
⑥¶디플로포리타강(Diploporita) : 오르도비스기 전기~데본기 전기(약 4억~5억 년 전).

바다나리아문(Crinozoa)
화석종과 현생종이 모두 있다(오르도비스기 전기[약 5억 년 전]부터 현재까지). 오방사대칭이며, 부드러운 부분이 5개 또는 더욱 복잡한 완이 있는 악부 속에 둘러싸여 있다.
바다나리강(Crinoidea)은 오르도비스기 전기(약 5억 년 전)부터 현재까지 있다.
병부가 있거나 2차적으로 없다. 악부는 작고 컵 모양으로 대개 가지를 치고 있으며, 먹이를 잡는 완에는 작은 우지가 매우 많다. 유병(有柄)인 바다술류와 무병(無柄)인 깃별나리류로 나뉘는 현존 관절아강(Articulata)이 있다. 약 700종의 현생종이 있다(→ 바다나리).

불가사리아문(Asterozoa)
화석종과 현생종이 있다(오르도비스기 전기[약 5억 년 전]부터 현재까지)(→ 불가사리아문). 5개의 축을 따라 완이 있어 별 모양의 몸을 갖는 방사대칭성이다.
중앙에 입이 있고, 5개의 완이 있으며 아래쪽에 관족과 입이 있다.
① 스텔레로이데아강(Stelleroidea) : 불가사리아문과 특징이 같다.
②¶소마스테로이데아강(Somasteroidea) : 오르도비스기 전기부터 약 3억 5,000만 년 전의 데본기 후기까지 있었다.
외관상 불가사리류와 같으나 관족이 자리잡을 홈이 없다.
③ 불가사리강(Asteroidea) : 화석종과 현생종이 있다(약 4억 6,000만 년 전의 오르도비스기 중기에서 현재까지). 약 1,800 현생종이 있다. 완은 넓고 속이 비어 있다. 길이로 된 성장축에 의해 완의 깃 모양 구조 또는 배열이 없어졌다. 관족은 수가 많고 몸의 구부(口部)에 홈을 따라 있다.
관족은 뾰족하거나 흡반이 있다. 호흡은 몸의 반(反)구면에 있는 아가미에 의해 이루어지기도 한다. 현존하는 것들로 유극목(spinulosa)·차극목(Forcipulata) 등이 있다(→ 불가사리).
④ 거미불가사리강(Ophiuroidea) : 화석종과 현생종이 있다(오르도비스기인 약 4억 6,000만 년 전부터 현재까지). 반은 완과 뚜렷이 구분된다. 완은 길고 가늘며 단단하다. 관족을 위한 홈이 없다. 흡관성 관족은 없다. 항문과 차극이 없다. 호흡은 몸의 구부면에 있는 아가미를 통해 이루어진다.
현존하는 것은 혁사미목(Phrynophiurida)과 폐사미목(Ophiurida) 등이 있다. 약 2,000종이 현존한다(→ 거미불가사리).
⑤ 콘켄트리키클로이데아강(Concentricycloidea) : 몸은 완이 없이 편평하고 반(盤) 모양이다. 수관계는 관족과 함께 구부면에 있다. 수관은 이중(二重)환으로 이루어져 있다. 페리포디다목(Peripodida)이 있다. 2종이 현존한다. 1821년 이래로 명명된 현존 극피동물 가운데 최근의 새로운 강으로, 1986년에 알려졌다.
불가사리류로서, 단지 형태가 극단적인 것이라고 보는 사람도 있다.

성게아문(Echinozoa)
화석종과 현생종이 있다(약 5억 7,000만 년 전 캄브리아기 전기부터 현재까지). 근본적으로는 공 모양이며 2차적으로 원통형 또는 원반형의 몸을 가지는 방사대칭성 동물이다.
완과 우지 등은 없다.
①¶키클로키스토이데아강(Cyclocystoidea) : 오르도비스기 중기~데본기 중기(약 3억 7,500만~4억 6,000만 년 전).
②¶에드리오아스테로이데아강(Edrioasteroidea) : 캄브리아기 전기~석탄기 전기(약 3억 4,000만~5억 7,000만 년 전).
③¶에드리오블라스토이데아강(Edrioblastoidea) : 오르도비스기 중기(약 3억 7,000만 년 전).
④¶헬리코플라코이데아강(Helicoplacoidea) : 캄브리아기 전기(약 5억 7,000만 년 전).
⑤¶오피오키스티오이데아강(Ophiocistioidea) : 오르도비스기 전기~실루리아기 후기(약 3억 9,500만~5억 년 전).
⑥ 해삼강(Holothuroidea) : 화석종과 현생종이 있다(오르도비스기인 약 4억 6,000만 년 전부터 현재까지). 긴 원통형으로 한쪽 끝 또는 그 가까이에 입이 있고, 다른 한쪽 끝 또는 그 가까이에 항문이 있다.
입은 먹이를 먹을 때 사용하며 눈에 잘 띄는 촉수들이 이룬 환에 둘러싸여 있다. 극과 차극이 없으며, 단일 생식소이다. 골격은 현미경적 크기의 골편으로 축소되었다. 현존하는 것으로는 수수목(Dendrochirotida)·순수목(Aspidochirotida)·판족목(Elasipodida)·은종목(Molpadida)·무족목(Apodida) 등이 있다. 1,100여 종이 현존한다(→ 해삼).
⑦ 성게강(Echinoidea) : 화석종과 현생종이 있다(오르도비스기인 약 4억 6,000만 년 전부터 현재까지). 몸은 공 모양, 원반 모양, 알 모양이다.
움직일 수 있는 극과 차극을 지닌 골격으로 이루어졌다. 입은 아래쪽으로 향한다. 항문은 없다. 생식소는 5개 이하이다. 현존 관극목 등이 포함된 정형성게아강과 연잎성게목·염통성게목이 포함되어 있는 부정형성게아강으로 나뉜다. 현생종은 900여 가지이다.

한국의 극피동물
한국의 극피동물의 분류는 1879년 외국인 학자에 의해 시작되었으며 그후 1966년에 와서야 우리 학자에 의한 연구가 진행되었다.
지금까지 알려진 한국산 극피동물은 불가사리류 3목 15과 26속 41종, 거미불가사리류 2목 4아목 10과 23속 54종, 성게류 6목 13과 22속 27종, 해삼류 3목 6과 6속 13종으로 이들은 주로 조간대와 100m 이내의 아조대(亞潮帶)에서 채집한 표본이 대부분이다.
극피동물은 원래 좌우대칭동물군(Bilateria)에 속하나, 변태과정에서 2차적으로 방사대칭이 되어 오방사대칭을 보이는 후구동물(後口動物 Deuterostomia)이다. 또한 단체성이며 해산에만 있는 저서동물(benthos)로, 모두 해저에 살고 있다. 바다나리류 가운데는 고착생활을 하는 것이 있으나, 그밖의 극피동물은 바위나 모래진흙 속에서 움직인다.
한국산 극피동물의 지리적 분포상태를 보기 위해 여러 해양학적 환경조건을 고려, 한국해역을 동해·남해·제주도·황해 해역으로 나누어 해역별 분포상을 비교해보면 분류군의 생태적인 특성에 따라 차이는 있으나, 다음과 같다.
불가사리류의 경우 동해에 25종, 남해(제주도 포함)에 2종, 황해에는 5종이 있다.
거미불가사리류는 동해에 19종, 남해에 31종, 제주도에 23종, 황해에 8종이 분포하고 있다. 이와 같은 사실로 미루어볼 때 한국산 불가사리류와 거미불가사리류는 많은 종이 해안선의 굴곡이 심하고 섬이 많은 남해에 있으며, 수심이 얕고 주로 모래진흙 지대로 이루어진 황해에는 많지 않음을 알 수 있다.
각 해역에만 있는 종과 해역간의 공통 종을 살펴보면, 불가사리류는 동해와 남해에만 분포하는 종이 각각 12종이고 황해에만 분포하는 종은 없다.
또 공통으로 분포하는 종은 동해와 남해에 8종, 남해와 황해에 1종이 있고, 동해와 황해에만 공통으로 분포하는 종은 없으나 3면에 공통으로 출현하는 종은 4종이 있는 것으로 나타났다. 거미불가사리류는 54종 가운데 제주도에만 분포하는 종이 13종으로 가장 많고, 그 다음이 남해(대한해협) 11종, 동해 9종 순이며, 황해에는 1종만 분포하는 것으로 나타났다.
대양별 공유상태를 보면 불가사리류는 북태평양에도 분포하는 종이 14종으로 가장 많고, 그 다음이 인도·서태평양의 8종, 북극해의 7종, 북대서양의 4종 순이다.
거미불가사리의 경우에는 북태평양에 분포하는 종이 50종으로 가장 많고 그 다음 인도·서태평양이 20종, 북극해가 5종, 대서양이 5종 순이다. 이와 같이 우리나라가 위치한 북태평양에 가장 많은 종이 분포하고 있다. 해류의 영향을 받는 것으로 알려진 수형별 분포상태를 보면 거미불가사리의 경우, 한국이 온대에 위치해 있으므로 난온수형종이 가장 많고 그 다음이 남방에서 북상하는 쿠로시오 난류와 오호츠크 해로부터 남하하는 리만 해류의 영향을 받아 열대·난온수형종과 냉온종, 냉수형종의 순이다.
우리나라 연안에서 흔히 볼 수 있는 종을 극피동물의 종류별로 살펴보면 불가사리류의 경우, 현대목(Phanerozonia) 연변아목(Valvata) 선불가사리과(Linckiidae)의 빨강불가사리(Certonardoa semiregularis)가 제주도의 아조대에서 쉽게 볼 수 있는 종이며, 소주아목(Paxillosa) 검은띠불가사리과(Luidiidae)의 검은띠불가사리(Luidia quinaria)는 3면 연안의 아조대에서 흔히 보이고, 유극목(Spinulosa) 별불가사리과(Asrerinidae)의 별불가사리(Asterina pectinifera)는 3면 연안의 조간대에서 흔히 볼 수 있는 대표적인 종이다.
차극목(Forcipalata) 불가사리과(Asteriidae)의 문어다리불가사리(Plazaster borealis)는 완이 약 35개에 동해안의 아조대에서 채집되며, 아무르불가사리(Asterias amurensis)는 3면 연안의 아조대에서 흔히 볼 수 있는 대표적인 불가사리이다. 거미불가사리로는 혁사미목(Phrynophiurida) 삼천발이과(Gorgonocephalidae) 가지거미불가사리속(Astrocladus) 종류가 많이 분포하는데 특히 혹가시불가사리(Astrocladus coniferus coniferus)는 3면 연안에 흔히 있는 대표적인 종이다.
폐사미목(Ophiurida)에 속하는 종으로는 악사미아목(Gnathophiurina) 가시거미불가사리과(Ophiothricidae)의 짧은가시거미불가사리(Ophiothrix exigua)와 순사미아목(Chilophiurina) 빗살거미불가사리과(Ophiuridae)의 왜곱슬거미불가사리(Ophioplocus japonicus)가 우리나라 조간대의 자갈·암석 밑이나 틈에서 쉽게 채집되며, 살시빗거미불가사리(Ophiura sarsii)는 3면 연안의 아조대 모래진흙 지대에 흔하게 살고 있는 대표적인 종이다.
성게류 중에서는 정형성게류(Regularia) 성게목(Echinoida) 둥근성게과(Strongylocentrotidae)의 말똥성게(Hemicentrotus pulcherrimus)가 3면 연안의 조간대 바위 밑과 틈 사이에서 쉽게 볼 수 있는 대표적인 종이며, 동해와 남해 연안의 아조대에 있는 둥근성게(Strongylocentrotus nudus)와 보라성게과(Echinometridae) 보라성게(Anthocidaris crassispina)는 알을 식용으로 쓰는 중요한 수산자원으로, 해녀의 주된 작업대상이다.
부정형성게류(Irregularia)로서 연잎성게목(Clypeastroida) 쟁반연잎성게과(Scutellidae)의 연잎성게(Scaphechinus mirabilis)는 상주 해수욕장 등 남해 연안에 있는 해수욕장의 모래 밑에 살고 있으며, 모양이 이상하여 옛날부터 잘 알려져 있는 균열염통성게과(Schizasteridae)의 염통성게(Schizaster lacunosus)는 남해와 황해에서 10~20m 정도 깊이의 모래진흙 속에 묻혀 살고 있다.
해삼류는 순수목(Aspidochirota) 돌기해삼과(Stichopidae)의 돌기해삼(Stichopus japonicus)이 남해안에 흔히 서식하는 대표적인 종으로, 육질부분이 많아 식용으로 쓰이는 중요 수산자원이다.
수수목(Dendrochirotida) 광삼과(Cucumariidae)에 속하는 여러 광삼류(Cucumaria)는 조간대와 아조대의 바위 밑과 틈, 그리고 죽은 산호충류 등의 골축 사이에서 살고 있으며, 잎사귀해삼과(Phyllophoridae)의 보라해삼붙이(Afrocucumis africana)는 제주도 서귀포의 조간대에서 채집되는 종이다.
대개 1,000m 이하의 심해에서 볼 수 있는 판족목(Elasipoda)은 우리나라에서는 아직 보고되지 않았으며, 은족목(Molpadonia)의 고구마해삼(Molpadia musculus)은 동해안에서 어선의 어망에 걸려나오기도 한다. 무족목(Apoda) 닻해삼과(Synaptida)의 가시닻해삼(Protankgra bidentatos)은 남해와 황해의 조간대, 물이 빠진 간석지의 모래진흙 속에서 채집되는 대표적인 종이다.